亚洲色图 欧美色图 生境异质性及源株密度对互花米草入侵力的影响
发布日期:2024-12-03 19:59 点击次数:74

河口湿地位于海陆接壤处[1], 在净化环境、提供生物栖息地、保护生物种种性、认知海岸、保管区域生态均衡、调遣碳轮回等方面施展着不行替代的伏击作用[2-3], 是单元面积上生态管事价值最高的生态系统类型[4]。受陆海物资交织、泥沙输移、冲淤演变、径流和潮汐相互作用的影响, 河口湿地具有私有的当然条款和异质生境[5], 千里积物粒径、盐度、氧化还原电位、水能源等与潮汐密切关联的系列环境因子皆通常阐扬出高度的空间异质性[6]。关联河口湿地生境异质性与盐沼植物滋长和分散的讨论标明亚洲色图 欧美色图, 生境条款的异质分散大大加多了盐沼植物接管和诈欺资源的难度, 严重影响其滋长、繁衍和扩散[7-8]。
当然植物种群中普遍存在密度效应, 它对植物个体滋长及种群动态的调遣历程是动态的[9-10]。为了更好地适合不同生境类型, 克隆植物可通过密度调控保管个体及种群认知, 即跟着源株密度及种内竞争的转变, 植株存活率、克隆构型特征以及种群动态等方面均会产生变化[11-13]。王永健和钟章成[14]开展的蝴蝶花(Iris japonica)源株密度的控制实验标明, 不同源株密度对蝴蝶花滋长、繁衍及生物量分派有显赫影响。Worm和Beusch[15]的讨论标明, 运转密度对鳗草(Zostera marina L.)幼苗滋长和斑块面积扩大均有明显的正效应。黎磊等[10]讨论也标明, 密度效应主要通过影响邻株植物间地上和地下器官的资源竞争强度来调遣个体的资源设置。密度效应在河口湿地盐沼植物群落也较为明显, 举例, 随源株密度加多, 海三棱藨草(Scirpus mariqueter)和藨草(Scirpus triqueter)的单株平均生物量减少, 而藨草和糙叶薹草的总生物量加多, 三种莎草科植物存在明显的克隆变异, 阐扬出明显的密度依赖性及较强的形态可塑性[16]。
互花米草(Spartina alterniflora)附庸禾本科米草属(Spartina, 别名绳草属), 具有广大的克隆整合与环境饰变才智[17-18], 能在热带、亚热带到温带沿海地区广博的局面带分散, 频繁在河口、海湾等沿海滩涂的潮间带形成单优群落[19], 进而破裂生态系统结构和功能, 威胁生物种种性, 是典型的东说念主为引种后扩散蔓延形成生态危害的外来入侵物种[20]。现在, 互花米草入侵机制的讨论已等闲开展, 可详细为互花米草的告捷入侵取决于互花米草的入侵力、合适的入侵道路以及受体系统的可入侵性[21]。互花米草极强的入侵力与其生物学特质密切关联, 包括对异质生境的广适性、种种的繁育对策、广大的扩散与竞争才智[17, 22-23]。现在, 已围绕淹水[23]、盐度[24]、潮滩冲淤[25]、温度[26]、泥土水分[27]等环境条款, 开展了互花米草对异质生境的适合性讨论。然则, 关联互花米草的入侵力是否会受密度调控, 以及在生境异质性及源株密度共同影响下互花米草入侵力的变化尚有待久了讨论。
长江河口湿地是典型的异质潮滩, 受流域、河口与海域的多重压力的影响, 河口湿地生态系统更具多变性、脆弱性与敏锐性[28-30]。20世纪90年代, 互花米草被引种到长江口潮滩以积淤筑陆、保护河堤。经过假寓适合阶段后, 由于其极强的扩散与竞争才智, 现已在潮间带湿地苟且蔓延, 对腹地物种组成了严重威胁[31-32]。很多学者艰巨于长江口互花米草的入侵机制和防治战略的讨论[7-8, 33], 但关于异质性河口环境中互花米草生物学特质(举例密度)影响其入侵力的讨论尚不久了, 而互花米草入侵运转密度将可能决定其滋长与资源分派的相对高下[14], 枯竭对这一历程的了解也不利于互花米草入侵动态分析和优化控制本领建立。因此, 本讨论遴荐不同类型生境, 通过苦楚移栽实验, 模拟互花米草以不同运转密度入侵不同类型生境, 分析生境异质性及源株密度对互花米草假寓、滋长和扩散的影响, 久了探讨生境异质性及源株密度对互花米草入侵力的影响, 辨识影响互花米草入侵力的主要因子, 为我国开展互花米草防控及海岸带保护、建造和贬责提供科学依据, 具有伏击的表面和现实真理真理。
1 讨论区域与讨论体式
1.1 讨论区域概况
南汇东滩湿地(30°50'04"—31°06'47"N, 121°50'50"—121°51'40"E)位于长江河口南岸, 毗邻杭州湾(图 1), 是上海市潮滩淤长最快的湿地之一, 亦然典型的异质潮滩[34]。该区域地处北亚热带, 年平均气温15—16℃, 年平均降水量1022 mm, 局面温暖湿润, 属典型的季风局面[35]。潮流属于非正规半日浅波澜流, 水流呈走动清醒性质[36]。潮间带上风植物主要为芦苇(Phragmites australis)、海三棱藨草(S. mariqueter)和互花米草(S. alterniflora)。
1.2 实验体式及规划
1.2.1 实验规划
2016年4月, 在南汇东滩的当然潮滩缔造南(粉砂质潮滩)、北(淤泥质潮滩)两条断面, 隔离在每条断面的植被区及前沿光滩缔造固定实验样方, 其中S1生境中上景色种为海三棱藨草, S2生境为粉砂质光滩, N1生境中上景色种为芦苇, N2生境为淤泥质光滩(图 1)。诈欺及时动态定位系统(Real-Time Kinematic Global Position System, RTK-GPS)测得S1、S2、N1和N2生境的吴淞高程隔离为(2.7±0.12)、(2.3±0.21)、(3.0±0.29) m和(2.4±0.9) m, 均在互花米草可分散高程边界内[37]。4月10日, 在南汇东滩潮间带互花米草单优群落内, 挖取高度一致((18.28±1.65) cm)、长势健康的互花米草根茎苗, 并按1、3、5、10株/m2和20株/m2的源株密度赶紧移栽到固定样方内(1 m×1 m), 每个源株密度设3个重迭, 共60个移栽样方, 样方与样方之间隔绝2 m(图 1)。
1.2.2 植物监测
于总共滋长季(2016年4月10日—2016年10月10日)每月依期监测不同生境、不同源株密度处理下, 移栽互花米草根茎苗的存活率、密度及株高, 并计较种群增长率(公式1)。
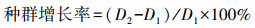
(1)
式中, D1为源株密度, D2为实验收尾时互花米草密度。
1.2.3 生境参数监测
植物监测同期, 经受象征桩法[38]监测各生境潮滩冲淤动态。在每个生境的移栽样方旁缔造一根冲淤象征桩, 每个生境15根, 共60根象征桩。象征桩长1.5 m, 垂直打入潮滩1 m控制以确保认知, 纪录象征桩败露在滩面以上的高度(M0)看成相对高程零点, 之后每月测量象征桩尖端向东、南、西、北30 cm处, 距滩面垂直距离的平均值(Mx), M0与Mx的差值即为该时段的冲淤变化, 恰恰为淤积, 负值示意侵蚀[39]。
2016年4月(盐沼植物存活重要时间), 按照“S”型赶紧多点羼杂的采样体式, 使用100 m3容积的环刀在每个样方内汇集3份重迭上层千里积物样品, 带回实验室后诈欺贝克曼库尔特Coulter LS13320型激光粒度仪进行粒度分析; 诈欺W.E.T泥土三参仪(英国Delta-T)现场原位监测泥土电导率; 诈欺RTK-GPS测得样带滩面高程数据后, 通过与国度海洋局发布的平均潮位数据(芦潮港站)进行耦合后, 取得不同生境的日均淹水时辰和相应的淹水深度。
1.3 数据处理
应用统计软件(IBM SPSS Statistics 19)对实验数据进行统计分析, 使用单因子方差分析(One-way ANOVA)中的最小显赫互异(LSD)对移栽后互花米草根茎苗的存活率、密度、株高及种群增长率进行互异显赫性测验(显赫性水平0.05);诈欺主因素分析体式对环境因子进行排序(旋转体式:最大方差法), 筛选出影响互花米草入侵力的重要环境因子。使用Origin 9.0软件作图。
2 成果
2.1 生境异质性及源株密度对互花米草存活的影响
不同处理下, 互花米草存活率均阐扬为跟着滋长季的延长, 先着过期趋于巩固的变化礼貌(图 2), 生境异质性及源株密度对互花米草存活率均有极显赫影响(P < 0.01, 表 1)。植被区(N1和S1)的互花米草存活率显赫高于前沿光滩(N2和S2)(P < 0.05, 图 2)。源株密度对互花米草存活的影响在不同生境有不同的阐扬。互花米草在植被区的存活率随源株密度加多而减小(图 2)。其中, 在N1生境(芦苇群落), 互花米草源株密度为1株/m2和3株/m2时, 其存活率均为100%;跟着源株密度的加多(5、10株/m2和20株/m2), 互花米草存活率略有着落, 但仍高于68%;在S1生境(海三棱藨草群落), 源株密度为1株/m2时, 互花米草存活率为100%;跟着源株密度加多(3、5、10株/m2和20株/m2), 互花米草存活率略有着落但均保管较高水平(>70%)。在光滩区(N2和S2), 除了源株密度为20株/m2的互花米草粗略一直存活在S2生境(粉砂质潮滩), 且能保留至滋长季末期外, 其余源株密度的互花米草根茎苗即使领先存活, 至滋长季末期也未能结束长期假寓(图 2)。
2.2 生境异质性及源株密度对互花米草株高的影响
滋长季末(10月), 异质生境及不同源株密度影响下互花米草的株高情状如图 3所示。生境异质性对互花米草株高有极显赫影响(P < 0.01, 表 1)。北断面植被区(芦苇群落, N1)互花米草平均株高为(148.86±9.99) cm, 显赫高于南断面植被区(海三棱藨草群落, S1)互花米草平均株高(74.7±8.87) cm(P < 0.05, 图 3)。此外, 在植被区, 跟着源株密度加多, 互花米草的株高无显赫互异(P > 0.05, 图 3)。光滩区惟一在S2生境, 且当互花米草源株密度为20株/m2时, 才能一直保捏存活, 滋长季末的株高为(43.33±5.35) cm, 显赫低于植被区(N1和S1)疏通源株密度处理(P < 0.05)。
2.3 生境异质性及源株密度对互花米草种群增长率的影响
生境异质性及源株密度均对互花米草种群增长率产生极显赫影响(P < 0.01, 表 1)。至滋长季末, 植被区(N1和S1)的互花米草种群全体呈现增长趋势(图 4), N1生境互花米草平均种群增长率((280.89±416.05)%)高于S1生境互花米草平均种群增长率((173.89±262.07)%); 而光滩(N2和S2)生境中, 互花米草种群全体则处于萎缩状态(图 4), N2生境由于移栽互花米草一齐被潮流冲走, 种群增长率为-100%, S2生境仅高源株密度(20株/m2)互花米草告捷假寓并扩散, 其种群增长率为(150±200)%, 显赫高于植被区(N1和S1)同源株密度处理(P < 0.05)。此外, 在植被区, 互花米草种群增长率与源株密度呈负关联(图 4)。其中, N1生境1株/m2源株密度处理下互花米草的种群增长率为(900±256.78)%, 显赫高于其余源株密度处理(3株/m2:(311.11±254.59)%; 5株/m2:(93.33±105.26)%; 10株/m2:(66.67±47.26)%; 20株/m2:(33.33±51.32)%)(P < 0.05);S2生境互花米草种群增长率变化趋势与N1相似, 即跟着源株密度加多, 种群增长率显赫裁减(P < 0.05, 图 4)。
2.4 不同生境环境因子分析
从表 2不错看出, 4个生境的环境因子存在明显互异。粒度分析成果标明, 北断面(N1和N2)中值粒径显赫小于南断面(S1和S2, P < 0.05, 表 2)。冲淤动态监测成果炫耀, 南断面(S1和S2)的滩面全体以淤积状态为主, 至11月, 植被区(S1)和光滩区(S2)滩面高程隔离累计抬升了(28.06±1.62) cm和(19.51±1.02) cm, 植被区的淤积速度显赫高于光滩(P < 0.05);而北断面(N1和N2)处于有冲有淤、全体微冲的状态, 植被区(N1)和光滩区(N2)滩面累计侵蚀量隔离为(0.55±3.28) cm和(5.99±1.72) cm, 光滩(N1)的侵蚀速度显赫高于植被区(N2, P < 0.05)。此外, 南北断面, 植被区泥土电导率、淹水时辰和淹水深度均显赫小于光滩(P < 0.05)。
环境因子主因素分析成果见表 3, 前2个特征值对应的累计方差孝顺率已达总方差的86.53%(>85%), 可代表所选想法的绝大部分信息。其中, 第一主因素占总方差51.80%, 该因素中淹水时辰和淹水深度载荷最高, 主要反应了高程对植物的影响。第二主因素与冲淤关联性最高, 主要阐扬千里积地貌对植物的影响。
3 盘考
3.1 密度效打法互花米草入侵力的影响
对克隆植物的讨论发现, 当生漫空间较充沛时, 高密度源株具有较强的存活才智及分蘖才智; 而当生漫空间有限时, 源株较多会导致竞争, 从而裁减植株存活率及分蘖才智[40-41]。本讨论中, 植被区由于已有腹地植物滋长, 移栽互花米草的生活空间相对有限, 密度效应明显。当源株密度越高时, 受到有限环境资源的控制和种间及种内竞争的影响也越大, 进而互花米草存活率和种群增长率也越低。这主如果由于植被区环境干扰小、生境认知, 在周围植物的遮护下, 以低源株密度定植的互花米草不错更好地诈欺有限资源, 生活并产生新分蘖, 相反, 高密度则会由于源株拥堵而导致竞争, 使无性分蘖及存活率着落[14, 16]。在光滩区, 由于不受生漫空间的控制, 高源株密度处理下互花米草存活率显赫高于低源株密度处理, 这与之前的互花米草移栽实验成果[7]也基本一致。这可能是因为光滩区域诚然有充沛的生漫空间, 但同期枯竭周围植物的卵翼, 高密度互花米草种群的“抱团”效应会促进附近源株的存活, 从而促进种群对生境的占领。赵春燕和沈有信[42]在模拟源株密度对入侵植物紫茎泽兰(Eupatorium coelestinum)滋长和竞争影响的讨论中也相通发现, 源株密度控制着紫茎泽兰叶片和根蘖株等构件和无性系植株的形成率, 源株密度越大, 紫茎泽兰个体株高、叶片数、根蘖株数和生物量越少, 而种群圆寂率越高。于国磊[43]开展的入侵克隆植物空腹莲子草(Alternanthera philoxeroides)种内关系的室外实验也得出了相似的论断, 即空腹莲子草的滋长和表型特征也受密度效应的调控, 其叶面积、节数、总生物量、根和叶生物量与密度呈显赫负关联。可见, 源株密度对克隆植物, 尤其是对以克隆繁衍为主要繁衍道路的外来物种的入侵力具有伏击的影响作用。
此外, 讨论也标明, 盐沼植物告捷假寓或扩散存在阈值效应, 包括生物阈值(生物量、根长等)[44-45]和环境阈值(高程、潮流、波浪等)[46-48]。在本讨论中, 互花米草的存活时辰总体与源株密度呈正关联。在植被区, 互花米草的存活和假寓不受源株密度控制, 1株/m2即可告捷入侵。在光滩区, 尽管该区域高程也曾粗略知足互花米草存活的最低高程要求, 但由于受潮流等环境因子的干扰相对较大, 互花米草存活的生物阈值也较高。仅当源株密度为20株/m2时, 互花米草不错告捷在粉砂质光滩(S2)假寓, 且可一直保留至曩昔滋长季末; 而在淤泥质光滩(N2), 当互花米草源株密度为20株/m2时, 在移栽初期和中期(4—8月)互花米草不错存活, 但跟着8月下旬的台风等环境条款的扰动, 最终未能保留至滋长季末, 可能需要更大的源株密度, 才能结束长期假寓。这也标明生境异质性也会影响互花米草入侵的生物阈值。
3.2 异质环境对互花米草入侵力的影响分析
外来入侵种的形态和生理特征通常会对异质生境阐扬出较强的可塑性, 其在不同资源和恫吓布景下的构型趋异是入侵才智的外皮特征体现[49-50]。表型可塑性被以为是展望外来种入侵力的一个特征[51]。许凯扬等[52]报说念不同生境(水生、湿生、中生和旱生)中的喜旱莲子草阐扬出显赫的表型趋异, 随泥土含水量减少, 喜旱莲子草的根冠比显赫加多, 分支强度显赫裁减。本讨论中, 高程越低、淹水恫吓越大、潮滩处于冲刷状态, 高源株密度处理下互花米草株高越低, 而种群增长率却越高, 阐扬出明显的表型趋异及对异质性生境的适合。这主要因为高程(或与其关联的环境因子)影响了互花米草的资源分派形状, 强恫吓条款下, 互花米草会优先蔓延种群, 缓解外界干扰压力, 晋升环境适合度, 进而告捷拓殖异质生境。沙晨燕等[53]在讨论典型滩涂环境因子对植物群幻灭间分散的影响时得出了相似的论断, 互花米草的盖度与密度随泥土紧实度、营养有用性增大而加多; 跟着高程的升高, 互花米草的生物量、株高反而着落。
生境条款的异质性大大加多了盐沼植物接管和诈欺资源的难度, 也严重影响其存活、繁衍和扩散[7-8]。本讨论中, 冲淤动态、淹水时辰和淹水深度是决定互花米草生活情状的主要影响因子。粉砂质光滩(S2)全体以淤积状态为主, 这在一定进程上促进了互花米草的假寓, 增强了已假寓植物对环境干扰的相悖力; 而淤泥质光滩(N2)滩面处于侵蚀状态, 千里积物的多量冲刷不利于假寓植物的保留。相干于千里积物淤积, 盐沼植物对侵蚀历程更为敏锐, 不同量级的千里积物扰动历程会径直或迤逦地影响植物存活[25]。此外, 在不同淹水时辰和淹水深度影响下, 互花米草的入侵力也阐扬出明显互异, 滋长季末90%以上的存活互花米草等分散在淹水时辰和淹水深度显赫较低的植被区(N1和S1), 其株高也显赫高于受淹水恫吓较大的光滩区存活植株。一方面可能是因为光滩区较长周期的潮流合并会导致植物缺氧、裁减光合营用, 径直抑制互花米草的滋长; 另一方面泥土氧化还原电位、海水矿物资、及光强等生境资源会因淹水时辰和深度增大而发生明显变化, 这可能也会迤逦影响互花米草的滋长和分散[54]。
3.3 河口湿地互花米草入侵力过火管控
入侵植物的入侵力取决于活力繁衍体的可传播性及定植契机窗口(一段可用来定植与繁衍的无干扰时辰)的时空可诈欺性[44, 55]。已有讨论标明, 互花米草有性繁衍体和无性繁衍体均可在潮流作用下进行远距离传播, 进而开荒腾达境[56-57]。盐沼植物种子随潮流传播运说念及实生苗定植契机窗口的讨论标明:受水能源及千里积能源的影响, 盐沼植物的种子很难通过长距离传播结束告捷假寓[58], 且即使告捷保留在种子库中的种子也并不料味着定植告捷[59]。对实生苗而言, 由于受到生态瓶颈(繁衍力、传播才智等)和物理压力(水能源、盐度等)阈值效应的影响及契机窗口的控制, 实生苗定植腾达境的告捷率普遍较低[60-61]。但在河口强潮能源作用下, 互花米草根茎苗或根状茎易被潮流冲刷并脱离母体[62], 诚然数目较少, 但假寓告捷率、滋长、繁衍等想法均优于有性繁衍状貌[18]。
国产拳交
本讨论中, 1株互花米草根茎苗即可在植被区结束认知存活、种群蔓延和告捷入侵, 当源株密度填塞大, 亦可告捷入侵光滩区。若不加以嗜好, 告捷假寓的零散互花米草能以无性繁衍的状貌充分占据空间上风, 结束鼎力蔓延的强健态势, 势必会对滩涂湿地的生态收复与生物种种性的保护带来极大的负面影响。此外, 连年来, 我国海岸带的圈围工程, 也显赫转变了潮滩的千里积环境, 导致泥沙在近岸区域捏续淤积, 并向海蔓延亚洲色图 欧美色图, 形成了大面积的腾达滩涂[63-64], 也为随潮流传播的互花米草繁衍体提供了合适的入侵生境。因此, 基于互花米草入侵机制的讨论, 在入侵早期, 通过合适东说念主为插手, 拦阻其进一步爆发, 可大大裁减互花米草的入侵风险和后期控制的经济资本, 对河口湿地的保育和可捏续贬责具有伏击真理真理。
